
What is a species?
Before we can authenticate "species", we have to know first, what "species" actually are. This is far from trivial.
Since the work by Mayr and Dobzhansky speciation is mainly described in terms of population genetics, as rise of propagation barreers that make genetically isolated subpopulations evolve into different directions. This concept was coined for speciation in animals, but can only partially be transferred to plant speciation, because genetic barreers are far more permeable in plants - hybridisations and even allopolyploidy is frequent in plants and render species more volatile as compared to animals. Concepts that are based upon solid and constant entities, will therefore not really work in plants. This meets population genetical, molecular, but also morphological criteria. A novel approach would be to consider "species" not as solid objects, but as processes that will culminate in different "species". This perspective shifts the focus on the mechanisms that let genetically linked populations, in a self-amplifying manner, split into subpopulations that will shift apart further, although the delineation is still permeable. Such mechanisms can be very subtle, even single mutations can cause speciation, when they channel sexuality (i.e. the gene flow between individuals) in one form or the other. "Channelling" means that the process which is triggered by this mutation, will launcht a positive feedback loop. One of the most basic channeling processes was the development of sexuality itself. In the very moment that sexuality initiated, this caused an irreversible change in the flow of evolution (Muller's Ratchet - the geneticist Muller used the ratchet as metaphor for a process that can only go on in one direction, because the teeth of the ratchet prevent that it can move backwards).
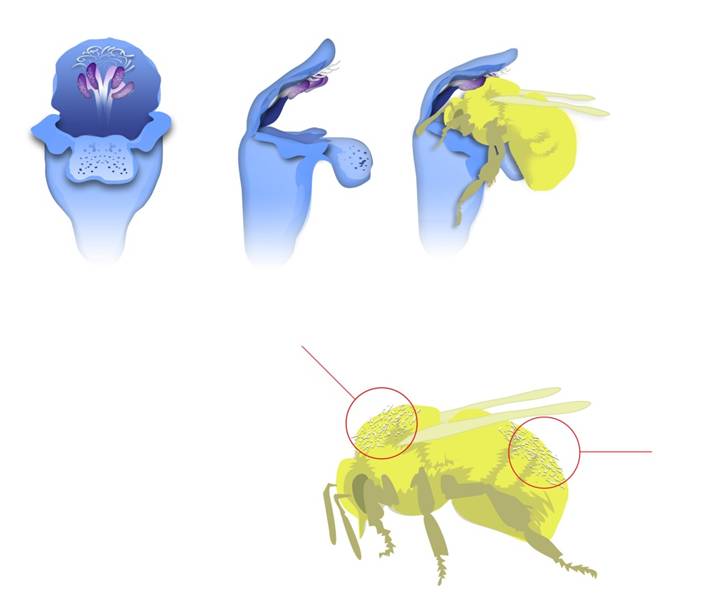
Model system: Channelling is very evident in traits linked with flower morphology. Even small morphological changes can shift gene flow in a massive manner, for instance, because other pollinators come into the play, or because the pollen is deposited on a different location on the pollinator. Famous examples are the lever mechanism of the Lamiaceae or heterostyly in primroses. To identify such "channelled" genes, one needs "non-channelled" references for comparison. Such references would be trait unrelated (most likely non-selected) genes, for instance microsatellites, introns or barcoding markers. Candidates for "channelled" genes would be factors linked with flower morphology. Here, detailed knowledge on the genetic control of flower development (EvoDevo) has become available during the last years. For instance, genes that regulate auxin transport in the stamina and thus the timing of anther growth would, upon mutation, cause a temporal isolation of the individual from the previous population or, possibly, a gene flow with a different plant species. Such a mutation would therefore amplify its own incidence and would qualify the respective locus as "gene of speciation".