
Microtubules and Stress in Rice
What is the topic?
Rice is the central staple food on this planet. At the same time, the flooded rice paddies are among the major players in the production of greenhouse gases on this planet. If we succeed to render rice more tolerant against drought, we can shorten the time, where paddies have to be flooded. Better drought tolerance therefore translates directly into better control of global warming. A second important topic is the optimisation of architecture - shorter, but more stable shoots mean reduced loss by lodging and windfall, better leaf angles mean better exploitation of solar radiation to build up yield.
Why are microtubules relevant here?
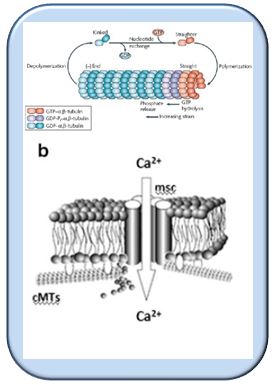
Microtubules impact all three traits - drought tolerance, shoot stunting, leaf angle. The orientation of so called cortical microtubules defines cell elongation or lateral expansion and, thus, the shape of a plant. In addition to this architectural function, microtubules are sensory in nature and perceive mechanical load (water scarcity, salinity stress, but also gravity). For this sensory function, it is not sufficient that microtubules are present - they have to assemble and disassemble as well. These dynamics determine, how plants can cope with adverse environmental conditions (stress). This provides approaches to improve stress tolerance. This is exactly, where we are heading to.
How do we address this topic?
Microtubules consist of two components, alpha- and beta-tubulin, which are genetically encoded in several variants. There exist two forms of alpha-tubulin - when the tubulin gene is expressed, first the full-length form is generated that harbours a tyrosine at the terminal position. This tyrosine can be cleaved off by an enzyme (the Tubulinyl-Tyrosin Carboxylase, TTC) yielding a detyrosinated alpha-tubulin. Interestingly, a different enzyme (the Tubulin-Tyrosin Ligase, TTL) can attach the tyrosine again. This generates a circuit of detyrosinated and retyrosinated alpha-tubulin. Detyrosinated tubulin dominates in stable microtubules, whereas very dynamic microtubules mainly consist of retyrosinated tubulin. This curious cycle seems to be conserved all over the eukaryotes (all organisms with a nucleus), at least all alpha-tubulins show this tyrosin signature at their terminal position. If something is conserved over so many different organisms including plants, fungi, and animals, it must be important - however, we are far from understanding what is going on here.
We have therefore cloned the TTL from rice an overexpressed this gene either in tobacco cells or in rice plants. In other words: we have created a situation, where tyrosinated tubulin is more abundant. We pretend that microtubules have become more dynamic than normally.
Now, we want to know, how the plant interprets this pretended situation. From this we hope to understand the biological meaning of the detyrosination- / retyrosination cycle. Once we have understood this, we can try in the next step to modulate this cycle by breeding in a way that is favourable for stress tolerance. Since we are dealing here with a signal and its interpretation, we hope that this will deliver very precise tools to adapt stress tolerance.